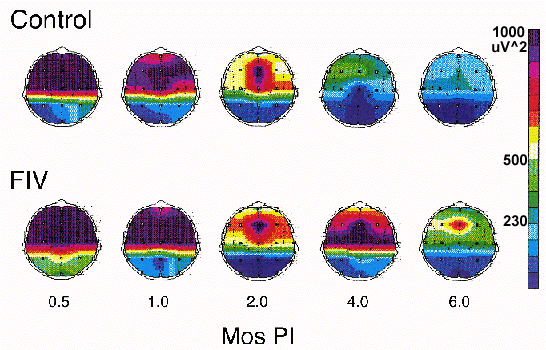| NeuroAIDS Vol. 1, No. 8, December 1998 |
| The feline model of neuroAIDS, part 1 (Part 2) |
| M. Podell,1,3 P. March,1,3 W. Buck,2,3 K. Hayes,2,3 M. Oglesbee,2,3 E. Steigerwald,1,3 and L. Mathes2,3 |
| 1Department of Veterinary Clinical Sciences, and 2Veterinary Biosciences, College of Veterinary Medicine |
| 3Center for Retrovirus Research, Comprehensive Cancer Center, The Ohio State University, 601 Tharp St., Columbus, Ohio 43210, United States |
| Address correspondence to: podell.1@osu.edu |
Abstract
![]() eline immunodeficiency virus (FIV) is a neurotropic lentivirus that produces a protracted state of immunodeficiency and encephalopathy in the cat. Recent evidence has shown several similarities to the natural progression of human immunodeficiency virus infection (HIV-1) associated degenerative effects on the central (CNS) and peripheral nervous system (PNS). This article will highlight the cumulative scientific body of evidence supporting the use of the feline model of neuroAIDS.
eline immunodeficiency virus (FIV) is a neurotropic lentivirus that produces a protracted state of immunodeficiency and encephalopathy in the cat. Recent evidence has shown several similarities to the natural progression of human immunodeficiency virus infection (HIV-1) associated degenerative effects on the central (CNS) and peripheral nervous system (PNS). This article will highlight the cumulative scientific body of evidence supporting the use of the feline model of neuroAIDS.
Introduction
Comparative virology
Although FIV and HIV-1 are relatively divergent at the genomic level, they share structural features common to all lentiviruses including gag, pol and env genes bounded by long terminal repeats, and several non-structural regulatory genes, notably vif and rev (1)(2). Functional equivalents of tat, vpu, vpr, and nef have not been demonstrated for FIV, although open reading frame A (ORF-A) may have limited transactivating activity (3). The FIV genome also encodes dUTPase, a common feature of ungulate, but not primate, lentiviruses, which may allow for increased genomic stability in vivo (4)(5).
Feline immunodeficiency virus has been subdivided into five clades (A-E) based on sequence similarities in the V3-V5 regions of env (6). Molecular clones of FIV have been used to evaluate: viral determinants of infection in vitro and in vivo (1)(5)(7)(8), target cell tropism (3), drug resistance (9), cytopathicity, genetic diversity and selection, neurovirulence (10), pathogenesis (11)(12), as well as for development of vaccines and potential gene therapy strategies (13)(14).
Temporal course of infectivity and immune responses
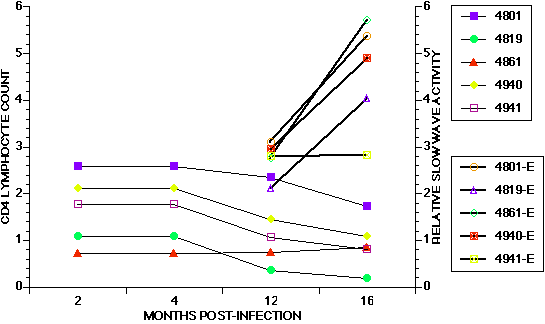
Of the animal lentivirus models for AIDS only the cat and monkey demonstrate parallel immunodeficiency with neurologic impairment after infection (15)(16)(17) (Figure 1).
|
|
Figure 1 (Enlarge): Temporal course of change in CD4 lymphocyte count and relative slow wave electroencephalographic activity (RSWA) in five adult FIV-MD infected cats. There is a progressive decline in CD4 lymphocyte count after 4 months post-infection that corresponds to a dramatic increase in RSWA in the majority of cats. This change demonstrate the progressive and inter-related nature of immunodeficiency and neurologic dysfunction in the feline model of neuroAIDS. |
Feline immunodeficiency virus is a naturally occurring lentivirus infection of cats, which causes an AIDS-like disease with neurologic sequelae remarkably similar in temporal expression, chronicity and severity to HIV-1 induced neuroAIDS (17)(18)(19)(20). Feline immunodeficiency virus infected cats undergo an acute phase of illness characterized by transient antigenemia, fever and lymphadenopathy. Commensurate with the decline in plasma antigenemia is the establishment of antiviral humoral and CTL responses (21). A prolonged asymptomatic phase then ensues during which cell-associated viremia persists (22) and CD4+ lymphocyte counts decline. One long-term study of cats experimentally infected with FIV observed 6/24 cats progressing to AIDS between 3.8 and 5.8 years PI (23). Frank immunodeficiency, increased viral burden, neurologic disease, opportunistic infections and neoplasia (19)(24) manifest end stage disease.
Like HIV-1, FIV is primarily cell-associated and lymphocyto-monocytotropic (25) with brain-derived isolates described as being primarily monocytotropic (26). Feline immunodeficiency virus and HIV-1 share the usage of at least one chemokine receptor (CXCR4) as a means to gain entry into target cells (27).
In vivo studies
Experimental infection studies of FIV neuropathogenesis
In nature, neurologic dysfunction resulting from FIV infection of the CNS is common and may be exacerbated by secondary infectious processes, such as feline infectious peritonitis (viral), toxoplasmosis (protozoal), and cryptococcosis (fungal) (24). The first report of neurologic effects of natural infection in cats was provide (18), with description of altered behavior in cats that died previously from a colony from which FIV was first isolated. Since then, most information about the effects and interaction of FIV on the nervous system have come from experimental inoculation In Vivo Studies that consistently demonstrate reproducible and measurable neurologic disease in the absence of secondary infection (16)(20)(28)(29)(30)(31) and others.
The first conclusive evidence that FIV is a neurotropic lentivirus was reported (16) where inoculation of adult cats with FIV-Petaluma via intracerebral, intra-bone marrow, and intravenous routes resulted in cerebrospinal fluid (CSF) pleocytosis and anti-FIV antibody production, and viral isolation from the subcortical brain structures. The CSF pleocytosis developed rapidly and persisted for a short duration of less than 20 weeks post-infection (PI), indicating a state of FIV-induced encephalitis. Development of anti-gag CSF antibodies occurred about 8 weeks after the appearance of the CSF pleocytosis in all cats. Thus, rapid infectivity of the brain was presumed to occur after peripheral inoculation, similar to HIV-1 infection.
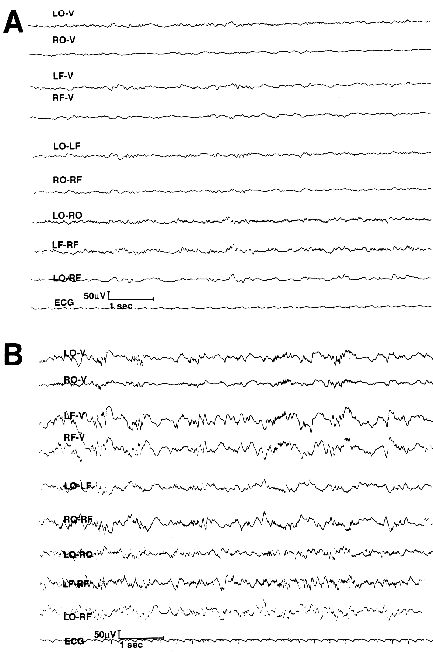
Several laboratories have confirmed the deleterious effect of FIV on neurologic function. Clinical and immunological evaluation was the first report on the onset and serial progression of immunodeficiency and encephalopathy (17). Intravenous FIV- Maryland (MD) challenged 8 week-old cats developed immunodeficiency, growth delay, and abnormal stereotypic behaviors as early as 4 weeks PI that persisted throughout the 16 month study. Serial neuroelectrodiagnostic evaluation revealed persistent abnormal electroencephalographic recordings (Figure 2) and prolonged latencies from visual and auditory evoked potential studies.
|
|
Figure 2 (Enlarge): Electroencephalographic tracings from a control (A) and FIV-MD 8 week-old infected cat (B) 12 months post-infection. The cats were 14 months old at the time of the recording which was done under general anesthesia with isoflurane. A bipolar left-over-right, front-to-back montage was used. Note the increased amplitude and frontally predominant polyomorphic delta wave dysrhythmia in the infected cat. LO=left occipital; RO=right occipital; LF=left frontal; RF=right frontal; ECG=electrocardiogram. (17). Reprinted with permission. Copyright (1993) Lippincott, Williams, and Wilkins. |
Virus was isolated from cortical and subcortical brain from all infected cats. Recent preliminary work in our laboratory has demonstrated a disruption in the normal cortical functional development in the FIV-MD infected kitten, with a significantly higher amount of slow wave activity at the 4 and 6 months PI time points (Figure 3).
|
|
Figure 3 (Enlarge): Temporal course of the relative change in topographic power spectral analysis of slow wave electroencephalographic activity (0.5-4.0 Hz) recorded from an age-matched uninfected and FIV-MD infected cat. Both cats were 8 weeks of age at the 0 time point when infection occurred. The FIV-MD infected cat has a significantly higher amount of slow wave activity at the 4 and 6 months (mos) post-infection (PI) time points. This change suggests a disruption in the normal cortical functional development in the FIV-MD infected animal. |
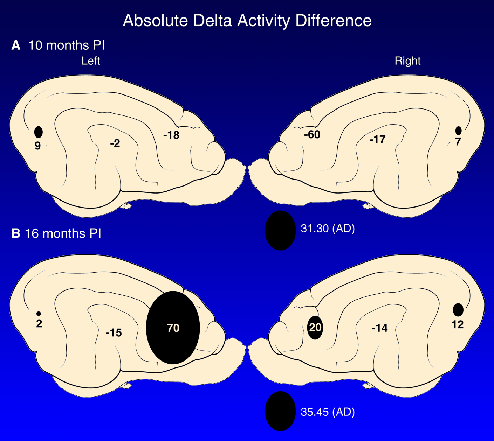
A comparative study of the effect of FIV-MD infection in adult cats revealed a delayed onset, progressive encephalopathy that paralleled the decline in the CD4/CD8 lymphocyte ratio (20). Specifically, the average amount of delta wave activity (extreme slow wave and therefore most abnormal activity) between FIV-MD and control cats was dramatically higher in the frontal cortex at 16 mo PI (Figure 4). These results indicated that age of infection influences the onset and severity of disease and may be associated with CD4 cell depletion in FIV-MD cats, as seen in HIV-1 infected people.
|
|
Figure 4 (Enlarge): Canograms of the difference between FIV-MD and control cats in the average absolute delta wave activity is presented. These plots demonstrate graphically the relationship between the amount and location of the slowest frequencies recorded (delta = 0.5 to 3.75 cycles per second) from all FIV-MD adult infected cats at 10 (A) and 16 (B) mo post-infection (PI). The more positive the value (AD; amplitude), the larger the canogram in the specific location, which translates into more delta wave activity for the FIV-MD group at that time point. A marked, preferential increase in average delta wave activity of the frontal cortex was found at 16 mo PI suggestive of a progressive worsening of cerebral function (20). Reprinted with permission. Copyright (1997) Lippincott, Williams, and Wilkins. |
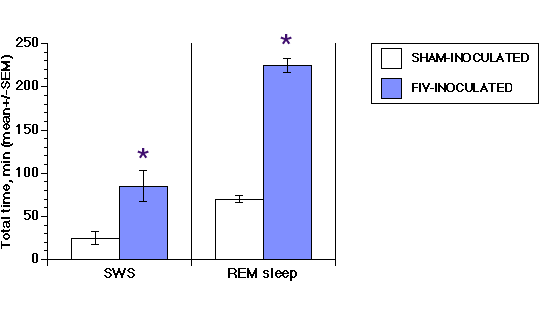
Working with the same FIV isolate, similar changes in challenged cats were described (30): abnormal neurologic examination and changes in sensory evoked potential associated with CD4 lymphocyte count depletion. Cerebrospinal fluid viral detection, viral antibodies, pleocytosis, and increased albumin leakage confirmed early infection of the CNS in both studies (17)(30). Further evidence of CNS infection and functional disruption was reported by Prospera-Garcia et al who demonstrated altered sleep architecture in FIV-MD infected cats. These infected cats spent 50% more time awake, had more sleep/waking stage shifts, and a 30% reduction of rapid-eye movement sleep than control cats (Figure 5). A similar situation was observed in rats inoculated intraventricularly with FIV-envelope protein, suggesting that this envelope glycoprotein may be responsible for changes in neurologic function (31).
|
|
Figure 5 (Enlarge): Delay in slow-wave sleep (SWS) and REM sleep onset is compared between FIV-infected and control cats. *, P=0.01. (32). Reprinted with permission. Copyright(1994) National Academy of Sciences, USA. |
An important development in studying the neuropathogenesis of FIV was the observation that an infectious molecular clone, pFIV-PPR, causes alterations in neurologic function (10). Infected cats developed abnormal gaits, changes in righting reflexes, behavioral changes, and alterations in sleep architecture. A knock-out gene mutant of this clone demonstrated that dUTPase was not essential for the neuropathogenesis. Preliminary data from our laboratory has demonstrated that inoculation of pFIV-PPR plasmid is pathogenic, with induction of CD4+ lymphocyte depletion and altered neurologic function (12). This infectious molecular clone allows the determination of the importance of specific viral elements in the neuropathogenesis of lentiviral infection.
Feline immunodeficiency virus also has a detrimental effect on the PNS. Reductions in peripheral motor and sensory nerve conductions occur after pFIV-PPR infections, similar to that reported for HIV-1 neuropathy (10). Recently, we have shown that FIV-MD induces an inflammatory polymyopathy in adult infected cats (33). Several similarities are apparent in comparison to HIV-1 associated polymyositis reported in humans, including perivascular and pericapillary lymphocytic infiltration, and myofiber necrosis, phagocytosis, and regeneration (Figure 6), and only CD8+ immunohistochemical phenotypic lymphocyte labeling.
|
|
Figure 6 (Enlarge): Fresh frozen gastrocnemius muscle biopsy section of a markedly affected muscle from an adult FIV-MD infected cat demonstrating the presence of necrotic fibers with loss of the intermyofibrillar membranous network, homogenous cytoplasm and invasion by mononuclear cells. Trichrome; Bar= 100 |
Evaluation of behavioral abnormalities with FIV infection has recently been given more attention in elucidating strain dependent neurovirulence. Neurobehavioral studies assessing open-field activity, plank walking performance, and development of blink reflexes demonstrated a differential neurovirulent effect of the isolate FIV-V1CSF compared to FIV-Petaluma (34). Kittens infected with FIV-V1CSF suffered from a delayed development of normal reflex activity, inability to navigate planks well, and reduced jumping ability. Our laboratory has demonstrated similar abnormalities in FIV-MD infected cats (35). The main difference demonstrated was a higher activity level and associated distractibility in FIV-infected cats that appears to be related to their overall deficient performance when learning new tasks. These results indicate that behavioral function is altered and cognition is quantitatively impaired in FIV-infected cats.
References
(1) Tomonaga K, Miyazawa T, Sakuragi J, Mori T, Adachi A, Mikami T (1993). The feline immunodeficiency virus ORF-A gene facilitates efficient viral replication in established T-cell lines and peripheral blood lymphocytes. J Virol 67(10):5889-95. Medline
(2) Phillips TR, Lamont C, Konings DA, Shacklett BL, Hamson CA, Luciw PA, Elder JH (1992). Identification of the Rev transactivation and Rev-responsive elements of feline immunodeficiency virus. J Virol 66(9):5464-71. Medline
(3) Waters AK, De Parseval AP, Lerner DL, Neil JC, Thompson FJ, Elder JH (1996). Influence of ORF2 on host cell tropism of feline immunodeficiency virus. Virology 1;215(1):10-6. Medline
(4) Elder JH, Lerner DL, Hasselkus-Light CS, Fontenot DJ, Hunter E, Luciw PA, Montelaro RC, Phillips TR (1992). Distinct subsets of retroviruses encode dUTPase. J Virol 66(3):1791-4. Medline
(5) Lerner DL, Wagaman PC, Phillips TR, Prospero-Garcia O, Henriksen SJ, Fox HS, Bloom FE, Elder JH (1995). Increased mutation frequency of feline immunodeficiency virus lacking functional deoxyuridine-triphosphatase. Proc Natl Acad Sci 1;92(16):7480-4. Medline
(6) Sodora DL, Courcelle J, Brojatsch J, Berson A, Wang YC, Dow SW, Hoover EA, Mullins JI (1995). Analysis of a feline immunodeficiency virus provirus reveals patterns of gene sequence conservation distinct from human immunodeficiency virus type 1. AIDS Res Hum Retroviruses 11(4):531-3. Medline
(7) Inoshima Y, Kohmoto M, Ikeda Y, Yamada H, Kawaguchi Y, Tomonaga K, Miyazawa T, Kai C, Umemura T, Mikami T (1996). Roles of the auxiliary genes and AP-1 binding site in the long terminal repeat of feline immunodeficiency virus in the early stage of infection in cats. J Virol 70(12):8518-26. Medline
(8) Pancino G, Castelot S, Sonigo P (1995). Differences in feline immunodeficiency virus host cell range correlate with envelope fusogenic properties. Virology 206(2):796-806. Medline
(9) North TW, LaCasse RA (1995). Testing anti-HIV drugs in the FIV model. Nat Med 1(5):410-1. Medline
(10) Phillips TR, Prospero-Garcia O, Wheeler DW, Wagaman PC, Lerner DL, Fox HS, Whalen LR, Bloom FE, Elder JH, Henriksen SJ (1996). Neurologic dysfunctions caused by a molecular clone of feline immunodeficiency virus, FIV-PPR. J Neurovirol 2(6):388-96. Medline
(11) Sparger EE, Beebe AM, Dua N, Himathongkam S, Elder JH, Torten M, Higgins J (1994). Infection of cats with molecularly cloned and biological isolates of the feline immunodeficiency virus. Virology 205(2):546-53. Medline
(12) Phillips A, Hayes K, Podell M, Buck W, Mathes L (1998). Paper in Preparation. J AIDS and Human Retrovirology
(13) Richardson J, Moraillon A, Baud S, Cuisinier AM, Sonigo P, Pancino G (1997). Enhancement of feline immunodeficiency virus (FIV) infection after DNA vaccination with the FIV envelope. J Virol 71(12):9640-9. Medline
(14) Cuisinier AM, Mallet V, Meyer A, Caldora C, Aubert A (1997). DNA vaccination using expression vectors carrying FIV structural genes induces immune response against feline immunodeficiency virus. Vaccine 15(10):1085-94. Medline
(15) Zink MC, Spelman JP, Robinson RB, Clements JE (1998). SIV infection of macaques--modeling the progression to AIDS dementia. J Neurovirol 4(3):249-59. Medline
(16) Dow SW, Poss ML, Hoover EA (1990). Feline immunodeficiency virus: a neurotropic lentivirus. J Acquir Immune Defic Syndr 3(7):658-68. Medline
(17) Podell M, Oglesbee M, Mathes L, Krakowka S, Olmstead R, Lafrado L (1993). AIDS-associated encephalopathy with experimental feline immunodeficiency virus infection. J Acquir Immune Defic Syndr 6(7):758-71. Medline
(18) Pedersen NC, Ho EW, Brown ML, Yamamoto JK (1987). Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science 235(4790):790-3. Medline
(19) Yamamoto JK, Sparger E, Ho EW, Andersen PR, O'Connor TP, Mandell CP, Lowenstine L, Munn R, Pedersen NC (1988). Pathogenesis of experimentally induced feline immunodeficiency virus infection in cats. Am J Vet Res 49(8):1246-58. Medline
(20) Podell M, Hayes K, Oglesbee M, Mathes L (1997). Progressive encephalopathy associated with CD4/CD8 inversion in adult FIV-infected cats. J Acquir Immune Defic Syndr Hum Retrovirol 1997 15(5):332-40. Medline
(21) Hayes KA, Lafrado LJ, Erickson JG, Marr JM, Mathes LE (1993). Prophylactic ZDV therapy prevents early viremia and lymphocyte decline but not primary infection in feline immunodeficiency virus-inoculated cats. J Acquir Immune Defic Syndr 6(2):127-34. Medline
(22) Diehl LJ, Mathiason-DuBard CK, O'Neil LL, Hoover EA (1995). Longitudinal assessment of feline immunodeficiency virus kinetics in plasma by use of a quantitative competitive reverse transcriptase PCR. J Virol 69(4):2328-32. Medline
(23) Mathiason-DuBard CK, Burkhard MH, O'Neil LL, Hoover EA (1998). Fourth International Feline Retrovirus Research Symposium, Glasgow, Scotland
(24) Sparger EE, Luciw PA, Elder JH, Yamamoto JK, Lowenstine LJ, Pedersen NC (1989). Feline immunodeficiency virus is a lentivirus associated with an AIDS-like disease in cats. AIDS 3 Suppl 1:S43-9. Medline
(25) Beebe AM, Faith TG, Sparger EE, Torten M, Pedersen NC, Dandekar S (1994). Evaluation of in vivo and in vitro interactions of feline immunodeficiency virus and feline leukemia virus. AIDS 8(7):873-8. Medline
(26) Dow SW, Dreitz MJ, Hoover EA (1992). Feline immunodeficiency virus neurotropism: evidence that astrocytes and microglia are the primary target cells. Vet Immunol Immunopathol 35(1-2):23-35. Medline
(27) Willett BJ, Picard L, Hosie MJ, Turner JD, Adema K, Clapham PR (1997). Shared usage of the chemokine receptor CXCR4 by the feline and human immunodeficiency viruses. J Virol 71(9):6407-15. Medline
(28) Podell M, Mathes L, Oglesbee M, hayes K, Beachy K (1995). Soc Neurosci 21, 236
(29) Podraza AM, Bornstein RA, Whitacre CC, Para MF, Fass RJ, Rice RR Jr, Nasrallah HA (1994).Neuropsychological performance and CD4 levels in HIV-1 asymptomatic infection. J Clin Exp Neuropsychol 16(5):777-83. Medline
(30) Phillips TR, Prospero-Garcia O, Puaoi DL, Lerner DL, Fox HS, Olmsted RA, Bloom FE, Henriksen SJ, Elder JH (1994). Neurological abnormalities associated with feline immunodeficiency virus infection. J Gen Virol 75 ( Pt 5):979-87. Medline
(31) Prospero-Garcia O, Herold N, Waters AK, Phillips TR, Elder JH, Henriksen SJ (1994). Intraventricular administration of a FIV-envelope protein induces sleep architecture changes in rats. Brain Res 659(1-2):254-8. Medline
(32) Prospero-Garcia O, Herold N, Phillips TR, Elder JH, Bloom FE, Henriksen SJ (1994). Sleep patterns are disturbed in cats infected with feline immunodeficiency virus. Proc Natl Acad Sci 91(26):12947-51. Medline
(33) Podell M, Chen E, Shelton D (1998). Feline immunodeficiency virus associated myopathy in the adult cat. Muscle Nerve 21(12):1680-5. Medline
(34) Power C, Buist R, Johnston JB, Del Bigio MR, Ni W, Dawood MR, Peeling J (1998). Neurovirulence in feline immunodeficiency virus-infected neonatal cats is viral strain specific and dependent on systemic immune suppression. J Virol 72(11):9109-15. Medline
(35) Steigerwald E, Sarter M, March P, Podell M (1999). Effects of feline immunodeficiency virus on cognition and behavioral function in cats. J Acquir Immune Defic Syndr Hum Retrovirol 20(5):411-9 Medline
| Copyright © 2001 by The American Association for the Advancement of Science |