|
|
|
| NeuroAIDS Vol. 1, No. 6, October 1998 |
| Role of viral proteins in HIV-1 neuropathogenesis with emphasis on Tat |
| A. Nath,1,2 J. D. Geiger,4 M. P. Mattson,3 D. S. K. Magnuson,5 M. Jones,2 and J. R. Berger1 |
| 1Departments of Neurology, 2Microbiology and Immunology, and 3Anatomy and Neurobiology,University of Kentucky Medical Center, Rm. L-445 Kentucky Clinic, Lexington, Kentucky 40536, United States |
| 4Department of Pharmacology and Therapeutics, University of Manitoba, Winnipeg, Manitoba, Canada |
| 5Department of Neurosurgery, University of Louisville, Louisville, Kentucky, United States |
| Address correspondence to: anath@pop.uky.edu |
Abstract
| ||||||||||||
![]() IV-1 infection is the most common CNS infection in the world and
the most common cause of dementia in people < 60 years of age in
North America. HIV-1 associated dementia (AIDS dementia complex) has
a poor prognosis and it is marked by deficits in motor control, cognition
and behavior (1). Neuropathological changes include
infiltration of mononuclear cells, glial cell activation and loss of
select populations of neurons (2)(3).
Although neuronal functions are adversely affected during HIV-1 infection,
direct infection of neurons with HIV-1 has only rarely been demonstrated
(4)(5). Neuronal dysfunction and neuronal
cell death by apoptosis and necrosis can be caused by neurotoxic viral
proteins and, thus, these proteins may represent important contributors
to HIV-1 neuropathogenesis (6). However, the presence
of most of these proteins in the brains of patients with HIV-1 infection
have yet to be demonstrated and, of those detected, their levels in
the extracellular compartment, where their neurotoxic actions are thought
to be mediated, remain to be determined. Therefore, in studies focused
on establishing the pathogenic mechanisms occurring as a consequence
of HIV-1 viral protein-mediated neurotoxicity and on their pharmacological
amelioration, the doses/concentrations of these neurotoxins relevant
to the human condition is a concern. Our position, espoused here, is
that Neuro-AIDS researchers should look past the question of doses and
concentrations, and instead address mechanistic questions.
IV-1 infection is the most common CNS infection in the world and
the most common cause of dementia in people < 60 years of age in
North America. HIV-1 associated dementia (AIDS dementia complex) has
a poor prognosis and it is marked by deficits in motor control, cognition
and behavior (1). Neuropathological changes include
infiltration of mononuclear cells, glial cell activation and loss of
select populations of neurons (2)(3).
Although neuronal functions are adversely affected during HIV-1 infection,
direct infection of neurons with HIV-1 has only rarely been demonstrated
(4)(5). Neuronal dysfunction and neuronal
cell death by apoptosis and necrosis can be caused by neurotoxic viral
proteins and, thus, these proteins may represent important contributors
to HIV-1 neuropathogenesis (6). However, the presence
of most of these proteins in the brains of patients with HIV-1 infection
have yet to be demonstrated and, of those detected, their levels in
the extracellular compartment, where their neurotoxic actions are thought
to be mediated, remain to be determined. Therefore, in studies focused
on establishing the pathogenic mechanisms occurring as a consequence
of HIV-1 viral protein-mediated neurotoxicity and on their pharmacological
amelioration, the doses/concentrations of these neurotoxins relevant
to the human condition is a concern. Our position, espoused here, is
that Neuro-AIDS researchers should look past the question of doses and
concentrations, and instead address mechanistic questions.
Introduction
Several HIV proteins have been shown to have neurotoxic properties in vitro. However, their presence in autopsy brain, particularly in extracellular compartments, has been difficult to demonstrate using conventional histopathological techniques. In this manuscript we examine the existing experimental evidence to help determine if these proteins play a role in the pathogenesis of HIV dementia.
Pathogenesis
HIV-1 proteins are neurotoxic
The HIV-1 proteins shown to be neurotoxic include the envelope glycoproteins gp120, gp160 and gp41, the non-structural protein Nef, the trans-activating gene regulatory protein Tat, a protein that regulates viral replication Rev, and another HIV-1 accessory protein vpr (6)(7). The existence of mRNA for these proteins has been well documented in autopsy brains of HIV-1 infected individuals (8)(9). At least gp41, a transmembrane protein expressed on the cell surface of infected cells, and Nef can be demonstrated to be present intracellularly in infected cells within the brain (10)(11). Since microglial cells and macrophages in the brain undergo a productive infection (12), one would argue that the other HIV proteins must also be present in the brain.
The HIV-1 viral coat glycoproteins gp41, gp120, and their precursor gp160 have all been shown to be neurotoxic (6). gp120, a protein involved in regulating viral entry and in determining viral tropism, is indirectly neurotoxic. Its actions are mediated via microglia and astrocytes (13), although some have suggested direct actions on neurons (14). The mechanisms implicated in gp120-induced neurotoxicity include decreased glutamate re-uptake by astrocytes, increased arachidonic acid release which then inhibits re-uptake of glutamate by neurons and astrocytes (15)(16), and increased levels of intracellular calcium in astrocytes and neurons (17). gp41 causes neurotoxicity in the presence of glial cells. Its mechanism of action includes nitric oxide formation (10). Attempts to detect gp120 protein in HIV-1 autopsy brain or in brains of transgenic mouse models of HIV-1 infection have not been successful (18) possibly due to technical factors (see below). But levels in the picomolar range have been demonstrated to result in neurotoxicity (19). Given the presence of gp120 mRNA, the well documented presence of gp41 protein (10), and the fact that both gp41 and gp120 are formed from gp160, the absence of gp120 in HIV-1 infected brain is highly unlikely. However, the duration of expression of gp120 will depend upon its rate of degradation which is yet to be determined.
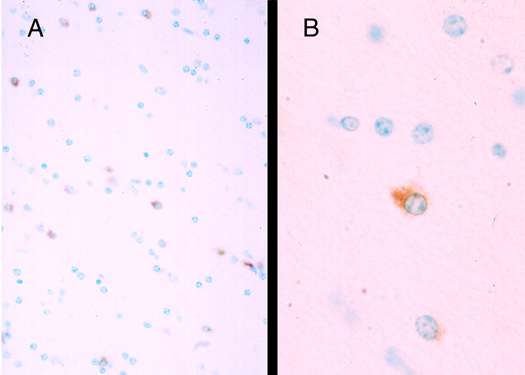
Recently, attention has shifted somewhat towards the study of the neurotoxic properties of Tat, a non-structural viral protein essential for viral replication. The first exon of Tat encodes for the initial 72 amino acids and the second exon forms the remaining 14 to 32 amino acids (20). Tat transcripts are elevated in HIV-1 demented brain (8) and both Tat1-72 (derived from the first exon) and Tat1-86 to 101 (full length) are produced by HIV-1 infected cells (21). Mechanisms implicated in the neurotoxic actions of Tat include direct depolarization of neurons, increased levels of intracellular calcium, increased production/release of pro-inflammatory cytokines, increased macrophage infiltration, activation of excitatory amino acid receptors by, as yet, uncharacterized mechanisms, and increased incidence of programmed cell death (apoptosis) (6)(22)(23). Despite, the documented presence of mRNA for Tat in HIV-1 autopsy brain (8)(9), investigators have questioned the potential relevance of findings on Tat neurotoxicity in the absence of evidence of Tat protein in HIV-1 infected brain. This position appears to be dramatically at odds with the well documented requirement of Tat protein for HIV-1 trans-activation. In other words, because the virus is replicating, Tat protein must be present. Here, we report preliminary findings that Tat protein, as demonstrated immunohistochemically, is present in HIV-1 infected human brain and in chimeric simian-human immunodeficiency virus infected brain of macaque monkey (Figure 1)(Nath et al., unpublished observations). These results show that Tat protein is present in close proximity to infected cells and neurons, and these findings bring us one step closer to determining the amounts of Tat protein present in HIV-1 infected tissues.
|
|
Figure 1 (Enlarge): Tat-immunoreactive cells in SHIV infected macaque. Sections from paraffin embedded tissue from the frontal white matter show cells immunostaining for Tat with rabbit polyclonal antisera raised against Tat1-72 and immunoaffinity purified. Sections were developed with diaminobenzidine to give a brown reaction product. (A) Several Tat positive glial cells are noted (magnification 40x). (B) Distinct cytoplasmic staining in a glial cell (magnification 100x). The neuropathological findings of this SHIV infected macaque (#44D) have been described previously (24). |
Neurotoxicity of other HIV-1 proteins has also been demonstrated. Peptides of HIV-1 Rev, when injected icv, were lethal (25) and in lymphoid cells caused necrotic cell death (26). Furthermore, Vpr was shown to cause neurotoxicity and caused membrane depolarization of neurons (7). Nef, an accessory protein that is over-expressed in HIV-1 infected astrocytes (27), and Nef-derived peptides may be neurotoxic (28). Although mRNA for these accessory proteins has been demonstrated in HIV-1 infected brain, protein expression has yet to be reported.
The role of each of these HIV-1 proteins in cell death and apoptosis in specific sub-populations of neurons remains uncertain. There are many factors that affect the levels of these HIV-1 proteins in brain and a discussion of these factors is important because of their obvious relevance to discussions of HIV-1 protein-induced neurotoxicity. These factors include, but are not necessarily limited to (because this field is relatively new and evolving quickly), the requirement for the continuous presence of the proteins, the in vitro and in vivo models used, the physiochemical properties of the viral proteins, and the size of the extracellular space which then dictates the concentrations reached in this compartment.
Continued presence of HIV-1 proteins may not be required for neurotoxicity: Evidence for a hit and run phenomenon
Toxic substances need not be continuously present to cause neurodegeneration. They only need to initiate a cascade of events that leads to neuronal degeneration. Glutamate, for example, applied to cells for brief intervals causes neuronal cell death hours later (29). For the HIV-1 protein Tat, we found in vitro that neurons responded with prolonged depolarizations in less than a second following Tat application (Figure 2)(30).
|
|
Figure 2 (Enlarge): Neuronal firing following a transient exposure to Tat: CA1neurons in hippocampal slices taken from 12 to 14 day old Sprague Dawley rats were exposed to recombinant Tat1-86 protein as previously described (30). A total of 5 pulses of ~400 fmoles each were delivered at a duration of 640 msec/pulse by pressure ejection thorough non-silanized glass pipettes. Exposure to Tat induces a rapid depolarization and action potentials. Recovery (repolarization) towards the resting membrane potential is slow and incomplete. |
Similarly brief (minutes) exposures to glial cells or macrophages leads to a delayed but prolonged release of cytokines (31). Following a single injection in vivo progressive pathological changes were observed over several days (32). Further, in a transgenic mouse model, where HIV proteins are expressed in oligodendrocytes at the end stage of spinal cord white mater disease, macrophages clear all markers of HIV presence in the cord (33). Similar studies with other HIV-1 proteins specifically addressing the length of exposure time necessary to initiate neurotoxicity need to be performed. Periodic release of small amounts of viral proteins may be sufficient to produce progressive neurodegeneration in a "hit and run " type of phenomenon. By focusing on the presence and quantity of viral proteins at the time of death the neurotoxic potential of these proteins will be underestimated. Indeed, in our in vivo studies, Tat could not be detected 6 hours post injection even though progressive neuropathological changes were noted 7 days later (32). By analogy, looking for viral proteins in autopsy brain tissues of HIV-1 infected patients may be similar to looking for a criminal at the site of a crime months after the crime was committed.
Limitations
In vitro assays must produce, in short defined intervals, measurable amounts of HIV-1 protein-induced neurotoxicity
HIV dementia and other neurodegenerative diseases progress gradually over months or years. Such long time intervals are not practical or feasible for in vitro experiments. Hence, in vitro conditions are purposely modified to accelerate, as well as enhance, responses to potentially toxic agents. Examples of such modifications include, excluding glia from neuronal cultures because of their ability to ‘buffer’ the toxins including released glutamate and glutamate-like substances, using selectively vulnerable cells like hippocampal neurons, and reducing concentrations of nutrients, growth factors and antioxidants in bathing media. Typically, dosages are used to achieve 30-50% neuronal cell death, against which neuroprotective effects of compounds can then be studied. One example of the consequence of modifying in vitro conditions comes from our studies of Tat neurotoxicity where 50% neuronal cell death was observed at 1 mM (30) in the presence of, but at 0.25 mM in the absence of serum and growth supplements (22). Further studies are necessary to determine if these viral proteins may have synergistic responses with one another or with other neurotoxic substances such as quinolinic acid and tumor necrosis factor which have been implicated in the pathogenesis of HIV dementia. Because HIV dementia is a chronic neurodegenerative process where small numbers of cells are likely dying over prolonged periods of time, doses of HIV-1 proteins smaller than those used in the laboratory would likely cause neurological dysfunction and neurodegeneration. Similarly, in Alzheimer's disease amyloid b peptide has been shown to be neurotoxic and thus implicated in the pathogenesis of Alzheimer's disease. However, the concentrations of 1 to 100 mg/ml used in vitro to study mechanisms of A b neurotoxicity are up to 10,000-times higher than A b levels in CSF of Alzheimer’s disease patients (34). Aged rhesus monkeys are susceptible to much lower concentrations of A b closer to those prtsent in Alzheimer's disease patients (35). Similar age-related susceptibility may be relevant to HIV dementia as well since older individuals are more susceptible to dementia with HIV infection. However, neurotoxicity assays in vitro are done using fetal or neonatal neurons, which may be more resistant to neurotoxicity. Despite the above short comings of in vitro experiments, they still represent powerful tools to describe neurodegenerative mechanisms that underlie HIV infection which can then be tested in more cumbersome and expensive in vivo models.
Physiochemical properties of HIV-1 viral proteins
Tat, gp120, and possibly other HIV-1 proteins, stick to glass and plastic. Such physiochemical properties makes working with these proteins challenging because plastic and glass vessels are used for the production and storage of the proteins as well as for determining their neurotoxic properties in vitro and in vivo. When vessels and syringes are silanized, it is not uncommon to observe a 10-fold decrease in the dose required to observe neurotoxicity in vivo (36). However, a similar approach for in vitro culture experiments is not possible because cultured cells will not attach and grow on "non stick " surfaces. This is why pressure-application of HIV-1 proteins loaded into micropipettes has proven to be an effective technique with which to measure, for example, Tat-induced depolarization, increases in levels of intracellular calcium, and neuronal cell death at doses as low as 2 to 20 femtomoles (37)(38).
The neurotoxic properties of gp120 and Tat are also highly dependent upon their tertiary configuration (39)(Holden et al., submitted). These proteins rapidly lose their neurotoxic potential when subjected to freeze-thaw conditions or if heated above physiological temperatures. For example, a single freeze-thaw cycle may result in a 50% reduction in Tat-induced neurotoxicity (30). The transactivation domain and the neurotoxic epitope of Tat contains 6 cysteine residues which are easily oxidized (39). This is also the most conserved region of Tat amongst all HIV strains (40). However, Tat looses its functional properties upon oxidation and because of this, Tat purification procedures require the presence of reducing agents and the final steps must be performed anaerobically. While these actions help maintain Tat stability, the presence of reducing agents would, almost certainly decrease its neurotoxicity by buffering oxygen free-radical induced cell death. Thus, physiochemical properties of Tat and gp120 significantly reduce bioavailability and these properties help explain some of the batch to batch variability that others and we have noted in determining dose-response profiles for neurotoxicity (Nath et al., unpublished observations; Amini and Alexander, Allegheny University, personal communication).
Extracellular space in the brain and viral protein concentrations
In general, animal viruses inefficiently incorporate structural proteins into viral particles, hence, cytopathic infections cause release of viral proteins into the extracellular space. These viral proteins then have the opportunity to interact with uninfected cells to cause cellular dysfunction or toxicity. Although all HIV-1 proteins have not been similarly studied, it is clear that Tat is actively released from infected microglial cells (41) and lymphoid cells (23)(42). Thus, for Tat at least, its distribution in the extracellular space would be expected to be highest in close vicinity to infected cells. The extracellular space in the brain comprises nearly 20% of the total brain volume and the size of the space varies between regions (e.g. extracellular space in the CA1 sub-region of the hippocampus is one-half that in layer VI of the sensorimotor cortex). Furthermore, the extracellular space is a dynamic space and increases in potassium following neuronal depolarization, decreases in pH, or alteration in sodium concentrations can lead to decreases in the size of the extracellular space by as much as 50% (43). Factors such as tortuosity and glial cell swelling can also impact upon extracellular volumes, and this is certainly relevant for patients with HIV infection, because astrocytosis and hypertrophy of astrocytes are early and prominent findings (44). The relative concentrations of neurotoxic substances can increase significantly without any increase in the total amount present in that area. Techniques that involve the use of tissue extracts for determining viral protein concentrations are likely to underestimate the relevant in vivo concentrations.
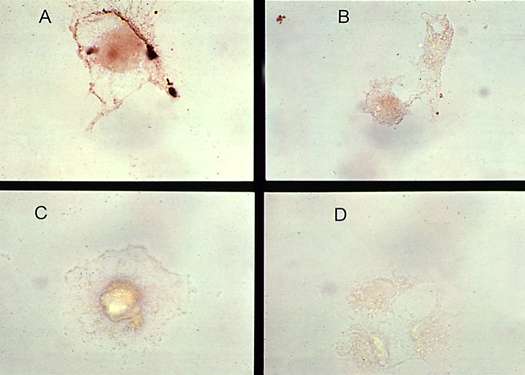
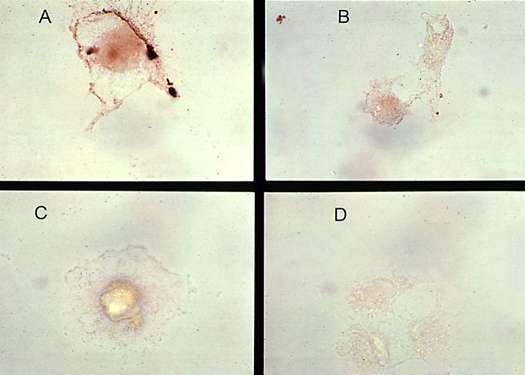
The half-life of HIV-1 proteins in extracellular spaces will affect their neurotoxic actions. Factors that control the half-lives of these substances include factors that regulate its release into the extracellular space, inactivation by proteases, and cellular transport. To date, only the HIV-1 protein Tat has been shown to be secreted from HIV-1 infected cells and this process has been characterized as being energy dependent (23). Once secreted, into the extracellular space, HIV-1 proteins may be released into the circulation, may bind to cell surface proteins on uninfected cells, or may be taken up into infected and uninfected cells. Both Tat and Vpr have been measured in the serum of HIV-infected patients (7)(45), both proteins bind to uninfected cells (7) (Figure 3), and, at least Tat, has been shown to be taken up by non-infected cells (46).
| Figure 3 (Enlarge): Tat binding to glial cells. Astrocytoid cells (U373 cells) were incubated with recombinant Tat1-72 protein for 2 hours in culture media, washed, post-fixed in 2% paraformaldehyde and blocked with 10% horse serum. The cells were then incubated with rabbit anti-Tat sera (1:1000) (A), or normal rabbit sera (B), followed by goat anti-rabbit sera conjugated to horse raddish peroxidase. Diaminobenzadine was used as a chromogen. (C) Tat treatment was omitted. (D) Rabbit anti-Tat sera was omitted. Distinct membrane staining is seen in panel A. |
The cellular uptake of Tat appears to be governed by the region of Tat protein formed by its second exon (46). Interestingly, neurotropic strains of HIV-1 from patients with HIV dementia show glutamate substitutions in the second exon (40) which is likely to decrease its ability to be taken up by cells, increasing its extracellular concentrations. Intracellularly, proteins such as the p6 gag protein bind to ubiquitin and are degraded by energy-dependent processes (47) but mechanisms for degradation of other HIV proteins need to be determined. Another mechanism by which HIV proteins may interact with surrounding uninfected cells is by cell-to-cell contact. For example, gp41 is a transmembrane protein that is expressed on the surface of infected cells which may induce neuronal injury in cells in close proximity (10).
Conclusions
We conclude that all viral proteins are present in the brain at one time or another. Their continued presence may not be necessary to cause neurotoxicity. A transient release of viral proteins may be sufficient to initiate a cascade of events that may last for prolonged periods of time. The in vivo concentrations of these proteins are dependent upon factors that regulate the production and release of the proteins in the extracellular space, the structural and physiochemical properties of the proteins themselves, and the dynamics of the extracellular space in the brain. In vitro assays invariably require large concentrations of proteins again due to their physiochemical properties and practical issues related to the design of these assays. The mechanisms responsible for neurotoxicity in AIDS is the fundamental issue; concentrations of HIV-1 proteins employed in in vitro assays are of secondary importance.
Acknowledgements
We would like to thank Dr. O. Narayan at the University of Kansas for providing us with tissues from the SHIV infected animals, and Drs. G. Alexander and S. Armini at Allegheny University for sharing with us their observations regarding physiochemical properties of Tat and for providing us with the protocol for silanizing glass. Martha Cooper assisted with preparation of figure 3. Dr. M. Ma performed the immunostaining in figure 4. Dr. JDG's research program is supported by grants from the National Health Research Development Program/Medical Research Council of Canada AIDS/HIV Research Strategy Program.
References
(1) Navia BA, Jordan BD, and Price RW (1986). The AIDS dementia complex: I. Clinical features. Ann Neurol 19, 517-524. Medline
(2) Masliah E, Ge N, Morey M, DeTeresa R, Terry RD, and Wiley C A (1992). Cortical dendritic pathology in human deficiency virus encephalitis. Lab Invest 66, 285-291. Medline
(3) Everall I, Luthert MBBS, Lantos MD (1993). A review of neuronal damage in human immunodeficiency virus infection: its assessment, possible mechanism and relationship to dementia. J Neuropathol Exp Neurol 52, 561-566. Medline
(4) Bagasra O, Lavi E, Bobroski L, Khalili K, Pestaner JP, Tawadros R, Pomerantz RJ (1996). Cellular reservoirs of HIV-1 in the central nervous system of infected individuals: identification by the combination of in situ polymerase chain reaction and immunohistochemistry. AIDS 10(6):573-85. Medline
(5) Nuovo GJ, Alfieri ML (1996). AIDS dementia is associated with massive, activated HIV-1 infection and concomitant expression of several cytokines. Mol Med 2(3):358-66. Medline
(6) Nath A, Geiger J (1998). Neurobiological Aspects of HIV infections: neurotoxic mechanisms. Prog Neurobiol 54(1):19-33. Medline
(7) Piller SC, Jans P, Gage PW, Jans DA (1998). Extracellular HIV-1 virus protein R causes a large inward current and cell death in cultured hippocampal neurons: implications for AIDS pathology. Proc Natl Acad Sci U S A 95(8):4595-600. Medline
(8) Wesselingh SL, Power C, Glass JD, Tyor WR, McArthur JC, Farber JM, Griffin JW, Griffin DE (1993). Intracerebral cytokine messenger RNA expression in acquired immunodeficiency syndrome dementia. Ann Neurol 33(6):576-82. Medline
(9) Wiley CA, Baldwin M, Achim CL (1996). Expression of regulatory and structural mRNA in the central nervous system. AIDS 10(8):843-7. Medline
(10) Adamson DC, Wildemann B, Sasaki M, Glass JD, McArthur JC, Christov VI, Dawson TM, Dawson VL (1996). Immunologic NO synthase: Elevation in severe AIDS dementia and induction by HIV-1 gp41. Science 274(5294):1917-21. Medline
(11) Saito Y, Sharer LR, Epstein LG, Michaels J, Mintz M, Louder M, Golding K, Cvetkovich TA, Blumberg BM (1994). Overexpression of nef as a marker for restricted HIV-1 infection of astrocytes in postmortem pediatric central nervous tissues. Neurology 44(3 Pt 1):474-81. Medline
(12) Gyorkey F, Melnick JL, Gyorkey P (1987). Human immunodeficiency virus in brain biopsies of patients with AIDS and progressive encephalopathy. J Infect Dis 155(5):870-6. Medline
(13) Lipton SA (1993). Human immunodeficiency virus-infected macrophages, gp120, and N-methyl-D-aspartate receptor-mediated neurotoxicity. Ann Neurol 33(2):227-8. Medline
(14) Lannuzel A, Barnier JV, Hery C, Huynh VT, Guibert B, Gray F, Vincent JD, Tardieu M (1997). Human immunodeficiency virus type 1 and its coat protein gp120 induce apoptosis and activate JNK and ERK mitogen-activated protein kinases in human neurons. Ann Neurol 42(6):847-56. Medline
(15) Lipton SA (1994). AIDS-related dementia and calcium homeostasis. Ann N Y Acad Sci 747:205-24. Medline
(16) Lipton SA, Yeh M, Dreyer EB (1994). Update on current models of HIV-related neuronal injury: platelet-activating factor, arachidonic acid and nitric oxide. Adv Neuroimmunol 4(3):181-8. Medline
(17) Nath A, Padua RA, Geiger JD (1995). HIV-1 coat protein gp120-induced increases in levels of intrasynaptosomal calcium. Brain Res 678(1-2):200-6. Medline
(18) Toggas SM, Masliah E, Rockenstein EM, Rall GF, Abraham CR, Mucke L (1994). Central nervous system damage produced by expression of the HIV-1 coat protein gp120 in transgenic mice. Nature 367(6459):188-93. Medline
(19) Dreyer EB, Kaiser PK, Offermann JT, Lipton SA (1990). HIV-1 coat protein neurotoxicity prevented by calcium channel antagonists. Science 248(4953):364-7. Medline
(20) Meyers G (1993). Human retroviruses and AIDS: A Compilation and Analysis of Nucleic Acids Sequences. Los Almos, New Mexico.
(21) Malim MH, Cullen BR (1991). HIV-1 structural gene expression requires the binding of multiple Rev monomers to the viral RRE: implications for HIV-1 latency. Cell 65(2):241-8. Medline
(22) New DR, Ma M, Epstein LG, Nath A, Gelbard HA (1997). Human immunodeficiency virus type 1 Tat protein induces death by apoptosis in primary human neuron cultures. J Neurovirol 3(2):168-73. Medline
(23) Chang HC, Samaniego F, Nair BC, Buonaguro L, Ensoli B (1997). HIV-1 tat protein exits from cells via a leaderless secretory pathway and binds to extracellelar matrix-associated heparan sulfate proteoglycan through its basic region. AIDS 11(12):1421-31. Medline
(24) Raghavan R, Stephens EB, Joag SV, Adany I, Pinson DM, Li Z, Jia F, Sahni M, Wang C, Leung K, Foresman L, Narayan O (1997). Neuropathogenesis of chimeric simian/human immunodeficiency virus infection in pig-tailed and rhesus macaques. Brain Pathol 7(3):851-61. Medline
(25) Mabrouk K, Van Rietschoten J, Vives E, Darbon H, Rochat H, Sabatier JM (1991). Lethal neurotoxicity in mice of the basic domains of HIV and SIV Rev proteins. Study of these regions by circular dichroism. FEBS Lett 289(1):13-7. Medline
(26) Nosaka T, Takamatsu T, Miyazaki Y, Sano K, Sato A, Kubota S, Sakurai M, Ariumi Y, Nakai M, Fujita S (1993). Cytotoxic activity of rev protein of human immunodeficiency virus type 1 by nucleolar dysfunction. Exp Cell Res 209(1):89-102. Medline
(27) Kohleisen B, Neumann M, Herrmann R, Brack-Werner R, Krohn KJ, Ovod V, Ranki A, Erfle V (1992). Cellular localization of Nef expressed in persistently HIV-1 infected low producer astrocytes. AIDS 6(12):1427-36. Medline
(28) Werner T, Ferroni S, Saermark T, Brack-Werner R, Banati RB, Mager R, Steinaa L, Kreutzberg GW, Erfle V (1991). HIV-1 Nef protein exhibits structural and functional similarity to scorpion peptides interacting with potassium channels. AIDS 5(11):1301-8. Medline
(29) Sucher NJ, Lipton SA, Dreyer EB (1997). Molecular basis of glutamate toxicity in retinal ganglion cells. Vision Res 37(24):3483-93. Medline
(30) Magnuson DS, Knudsen BE, Geiger JD, Brownstone RM, Nath A (1995). Human immunodeficiency virus type 1 tat activates non-N-methyl-D-aspartate excitatory amino acid receptors and causes neurotoxicity. Ann Neurol 37(3):373-80. Medline
(31) Nath A, Conant K, Chen P, Scott C, Major EO (1999. )Transient exposure to HIV-1 Tat protein results in cytokine production in macrophages and astrocytes: A hit and run phenomenon. J Biol Chem 274(24):17098-102. Medline
(32) Jones M, Olafson K, Del Bigio MR, Peeling J, Nath A (1998). Intraventricular injection of human immunodeficiency virus type 1 (HIV-1) Tat protein causes inflammation, gliosis, apoptosis, and ventricular enlargement. J Neuropathol Exp Neurol 57(6):563-70. Medline
(33) Goudreau G, Carpenter S, Beaulieu N, Jolicoeur P (1996). Vacuolar myelopathy in transgenic mice expressing human immunodeficiency virus type 1 proteins under the regulation of the myelin basic protein gene promoter. Nat Med 2(6):655-61. Medline
(34) Neve RL, Robakis NK (1998). Alzheimer's disease: a re-examination of the amyloid hypothesis. Trends Neurosci 21(1):15-9. Medline
(35) Geula C, Wu CK, Saroff D, Lorenzo A, Yuan M, Yankner BA (1998). Aging renders the brain vulnerable to amyloid beta-protein neurotoxicity. Nat Med 4(7):827-31. Medline
(36) Maragos W, Tillman P, Jones M, Nath A (1998). Pattern of hippocampal injury with HIV-1 Tat protein (abstract). J Neurovirology. 358
(37) Cheng J, Nath A, Knudsen B, Hochman S, Geiger JD, Ma M, Magnuson DS (1998). Neuronal excitatory properties of human immunodeficiency virus type 1 tat protein. Neuroscience 82(1):97-106. Medline
(38) Haughey NJ, Nath A, Holden CP, Geiger JD (1998). HIV-1 Tat activates calcium release from IP3 sensitive pools in human fetal neurons and astrocytes; neuroprotection results from signal block at the level of phospholipase C (abstract). J Neurovirology 353.
(39) Nath A, Psooy K, Martin C, Knudsen B, Magnuson DS, Haughey N, Geiger JD (1996). Identification of a human immunodeficiency virus type 1 Tat epitope that is neuroexcitatory and neurotoxic. J Virol 70(3):1475-80. Medline
(40) Bratanich AC, Liu C, McArthur JC, Fudyk T, Glass JD, Mittoo S, Klassen GA, Power C (1998). Brain-derived HIV-1 tat sequences from AIDS patients with dementia show increased molecular heterogeneity. J Neurovirol 4(4):387-93. Medline
(41) Tardieu M, Hery C, Peudenier S, Boespflug O, Montagnier L (1992). Human immunodeficiency virus type 1-infected monocytic cells can destroy human neural cells after cell-to-cell adhesion. Ann Neurol 32(1):11-7. Medline
(42) Ensoli B, Buonaguro L, Barillari G, Fiorelli V, Gendelman R, Morgan RA, Wingfield P, Gallo RC (1993). Release, uptake, and effects of extracellular human immunodeficiency virus type-1 Tat protein on cell growth and viral replication. J Virol 67(1):277-87. Medline
(43) Sykova E (1997). Extracellular space volume and geometry of the rat brain after ischemia and central injury. Adv Neurol 73:121-35. Medline
(44) Vitkovic L, da Cunha A (1995). Role for astrocytosis in HIV-1-associated dementia. Curr Top Microbiol Immunol 202:105-16. Medline
(45) Westendorp MO, Frank R, Ochsenbauer C, Stricker K, Dhein J, Walczak H, Debatin KM, Krammer PH (1995). Sensitization of T cells to CD95-mediated apoptosis by HIV-1 Tat and gp120. Nature 375(6531):497-500. Medline
(46) Ma M, Nath A (1997). Molecular determinants for cellular uptake of Tat protein of human immunodeficiency virus type 1 in brain cells. J Virol 71(3):2495-9. Medline
(47) Ott DE, Coren LV, Copeland TD, Kane BP, Johnson DG, Sowder RC 2nd, Yoshinaka Y, Oroszlan S, Arthur LO, Henderson LE (1998). Ubiquitin is covalently attached to the p6Gag proteins of human immunodeficiency virus type 1 and simian immunodeficiency virus and to the p12Gag protein of Moloney murine leukemia virus. J Virol 72(4):2962-8. Medline
McArthur, J.C., Hover, D.R., and Bacelar, H. (1993). Dementia in AIDS patients. Neurology 43, 2245-2252. Medline
| Copyright Information | Site map |

{kind=link}