|
|
|
| NeuroAIDS Vol. 1, No. 4, August 1998 |
| In vitro blood-brain barrier model for HIV-induced CNS disease |
| D. Janigro,1 L. Strelow,2 G. Grant,1 and J. A. Nelson2 |
| 1University of Washington School of Medicine, Department of Neurological Surgery, Harborview Medical Center Box 359914, 325 9th Avenue, Seattle, Washington 98104, United States; Phone: (206)-731 2342; FAX: (206)-731 8543 |
| 2Oregon Health Sciences University, Department of Molecular Microbiology and Immunology, L220, 3181 SW Sam Jackson Park Road, Portland, Oregon 97201, United States; Phone: (503) 494-7769; FAX: (503) 494-2441 |
| Address correspondence to: ciao@u.washington.edu or ciao@u.washington.edu |
Abstract
| ||||||||||||
![]() everal in vitro tissue culture systems have been developed to reproduce the key physical and biochemical properties of the intact blood-brain barrier (BBB). However, most of these systems lack essential features which are characteristic of the in vivo BBB. We recently developed a dynamic in vitro BBB (DIV-BBB) model consisting of endothelial cells grown in the lumen of hollow fibers (capillaries) with astrocytes grown on the outside (extra-capillary space [ECS]). Exposure to pulsatile flow of the endothelial cells in the hollow fibers in the context of astrocytes induces characteristic BBB properties in these cells, including high transendothelial resistance, negligible permeability to sucrose and stereo-specific uptake of amino acids. In this review we will examine some of the properties of the DIV-BBB model and discuss some of the applications for examining mechanisms of HIV-induced NeuroAIDS and other central nervous system (CNS) diseases.
everal in vitro tissue culture systems have been developed to reproduce the key physical and biochemical properties of the intact blood-brain barrier (BBB). However, most of these systems lack essential features which are characteristic of the in vivo BBB. We recently developed a dynamic in vitro BBB (DIV-BBB) model consisting of endothelial cells grown in the lumen of hollow fibers (capillaries) with astrocytes grown on the outside (extra-capillary space [ECS]). Exposure to pulsatile flow of the endothelial cells in the hollow fibers in the context of astrocytes induces characteristic BBB properties in these cells, including high transendothelial resistance, negligible permeability to sucrose and stereo-specific uptake of amino acids. In this review we will examine some of the properties of the DIV-BBB model and discuss some of the applications for examining mechanisms of HIV-induced NeuroAIDS and other central nervous system (CNS) diseases.
Introduction
Blood-brain barrier
The BBB maintains the homeostasis of the brain microenvironment. Morphologically, it consists of specialized brain microvascular endothelial cells (BMEC) which line the intraluminal portion of brain capillaries and are in tight contact with astrocytes. They form a relatively impermeable barrier between peripheral blood and blood-borne substance and the brain parenchyma. This barrier is characterized by: (i) interendothelial tight junctions, (ii) reduced or absent pinocytosis, (iii) selective, asymmetric permeability to physiological ions and (iv) presence of BBB-specific transporters (Bradbury, 1993; Stanness et al., 1997).
HIV-induced neuroAIDS
AIDS dementia complex and other CNS disorders (Table 1) involve BBB dysfunction (Rhodes, 1991; Petito and Cash, 1992).
Diffuse leakage of non-permeable serum proteins into the brain parenchyma of post-mortem AIDS brains was found which was distinct from focal breakdown due to tissue necrosis. This suggests that abnormal vascular permeability may be an important process in the development of HIV-induced CNS dysfunction. Infection of brain endothelial cells may cause perturbations in BBB function, allowing toxic substances to cross into the normally inaccessible CNS. Alternatively, HIV-infected cells may traffic through a damaged BBB and thereby introduce virus and/or viral proteins into the CNS.
The luminal position of BMEC ensures that peripheral virus must encounter these cells before entering the brain. HIV has been shown to infect BMEC in vivo (Wiley et al., 1986; Mankowski et al., 1994) and in vitro (Moses et al., 1993; Mankowski et al., 1994 possibly resulting in BBB perturbations via disruption of tight junctions or by an alteration of selective transport mechanisms. The subsequent enhanced vascular permeability could result in the entry into the CNS of neurotoxic cytokines, ions and metabolites from the periphery, which would contribute to neuronal dysfunction.
Alternatively, viral infection of BMEC may change cell-surface expression of adhesion molecules possibly mediating recruitment and extravasation of virus-infected macrophages. Evidence for this model comes from studies of simian immunodeficiency virus (SIV) infection in rhesus macaques (Sasseville et al., 1994). While AIDS dementia complex remains a devastating consequence of HIV infection, the mechanisms of HIV entry into the CNS and the subsequent alterations contributing to pathological and behavioral abnormalities remain unknown.
Requirements for BBB Disease Models
Irrespective of whether the BBB disruption is the main pathogenic factor or an inevitable consequence of a CNS disease itself, our understanding of the cellular mechanisms that led to the disruption of the BBB are limited. This situation is due in part to the lack of comprehensive models of BBB. Any such models must reproduce the salient features of in situ BBB (Table 2), while allowing for manipulations aimed at mimicking the disease process.
DIV-BBB Model
Background
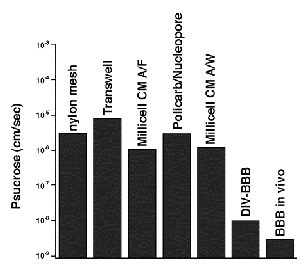
Currently several in vitro models of the BBB explore brain pathophysiology as well as the permeability and potential efficacy of drugs targeting the brain (Pardridge et al., 1990; Joo, 1985; Joo, 1993) (Figure 1).
| Figure 1. Comparison of the permeability values for different BBB models. |
The most commonly used substrate for culturing BMEC in vitro consists of a porous membrane support submerged in culture media. One such system, the bi-dimensional Transwell inserts (Costar), is characterized by a side-to-side diffusion. This system is capable of inducing polarity in the endothelial cells and affords an opportunity to study bi-directional transendothelial transport of solutes across the BBB (Pardridge et al., 1990; Joo, 1985; Joo, 1993). A major disadvantage of this system, however, is the lack of physiologic shear stress, which has been shown to induce tight junctional qualities in BMEC (Ott et al., 1995). Therefore, the tightness of the barrier in this type of model is typically much less stringent than that of in vivo BBB.
DIV-BBB model
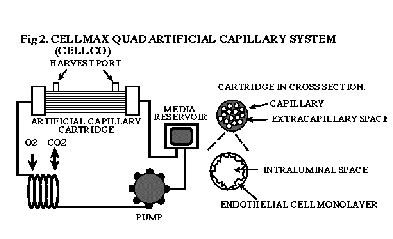
We have developed a dynamic, tri-dimensional in vitro culture system (DIV-BBB) which more closely mimics the in vivo BBB phenotype than other models (Figure 2).
| Figure 2. CELLMAX Quad artificial capillary system (Cellco). |
In this system, brain or peripheral endothelial cells are cultured in hollow fiber capillaries inside a sealed chamber (cartridge), with the endothelial cells grown intraluminally in the presence of astrocytes cultured abluminally. The hollow fiber cartridge system (Cellco/Spectrum) consists of artificial capillaries which are exposed to luminal pulsatile flow. When endothelial cells and astrocytes are cocultured, the DIV-BBB model is characterized by a high transendothelial electrical resistance (TER) of 1500-2000 Ohms/cm2, which is similar to that obtained in vivo. In addition to physiologic TER, the dynamic BBB model has successfully replicated several morphological and functional characteristics of the intact BBB. These include asymmetric potassium transport, stereo-selective transport of amino acids (L- vs. D- aspartate), expression of a BBB-like glucose transporter, the establishment of tight junctions and negligible permeability to radiolabelled sucrose (Stanness et al., 1996; Stanness et al., 1997).
Permeability of the BBB to CNS drugs
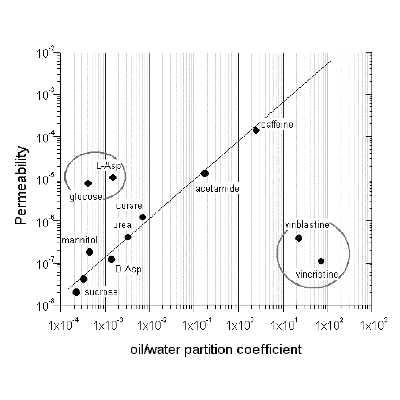
Apart from the exceptional and rare circumstance where drugs are actively transported directly into the CNS, the plasma-to-CNS concentration ratio is usually difficult to measure. Therapeutically relevant dosages in the CNS are difficult to achieve, since after oral or parenteral administration most drugs are excluded by the BBB. Rational CNS drug design cannot entirely and exclusively rely upon the physico-chemical properties of putative neurotherapeutics, since lipophilicity alone is a poor predictor for drug penetration into the CNS. This is particularly true for two large families of CNS drugs, antineoplastics and antivirals. Many important drugs and compounds fall quot;off the diagonalquot; relating lipophilicity to BBB permeability (Figure 3).
| Figure 3. Lipophilicity plot for various CNS compounds. |
This phenomenon is usually due to the presence of active transport systems. Therefore, a realistic requirement for testing CNS drugs in BBB models is that these systems must be capable of reproducing such transporters in vitro. The DIV-BBB model is capable of exhibiting such mechanisms of drug exclusion and transport.
Stereo-selective Transporters
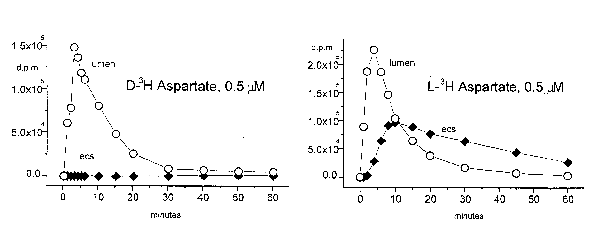
Further similarity between the selective drug permeability of the in vivo BBB and the DIV-BBB model has been found when comparing the permeability of non-BBB permeant amino acids that are transported across the BBB by a stereo-specific transporter. We found that the biologically active L-isomer of aspartic acid (L-Asp) was transported across the DIV-BBB at a significantly higher (>100) rate than its counterpart D-isomer (D-Asp) (Figure 4).
| Figure 4. Pharmacokinetic properties of the DIV-BBB model. L- or D-Aspartate was applied intraluminally and extraluminal samples taken at the intervals indicated. |
This is an important finding, because it suggests that this in vitro BBB model may be suitable for the study of drugs that are transporters. Note in Figure 4 the lack of extraluminal D-Asp accumulation compared to the passage of L-Asp under identical experimental conditions. The same DIV-BBB cartridge was used in both experiments.
Persistence of HIV and SIV in the DIV-BBB Model
In static cultures of HIV-infected BMEC, we could demonstrate virus persistence for the natural viable life of the culture. Cultures are viral DNA PCR positive and, while supernatant p24 values decrease to almost undetectable levels, infectious virus can be amplified from these supernatants by co-culture with CEMx174 indicator cells. Interestingly, cytopathic effect (CPE) in CEMx174 cells co-cultured with infected BMEC overnight before transfer to independent culture was far more rapid and profound than CPE in CEMx174 cells exposed only to infected BMEC supernatant.
These observations suggest that i) virus seeds from BMEC at a consistently low rate but transfer to CEMx174 cells is far more efficient in the context of cell-to-cell contact or that ii) the CEMx174 contact stimulus upregulates HIV replication in BMEC. These hypotheses need not be mutually exclusive. Importantly, these observations indicate that BMEC in a quiescent state are able to replicate HIV and that the virus can persist in these cells for an extended period of time. These are important observations for HIV-infection of BMEC in the DIV-BBB, since such culture systems can be maintained for extended periods of time and BMEC proliferation post-seeding is minimal.
Preliminary evidence accrued from studies using simian BMEC in the DIV-BBB model indicates that a persistent, long-term infection of BMEC can be established by the molecular clone of simian immunodeficiency virus strain SIVmac251. In brief, DIV-BBB cartridges were established with low passage primary simian BMEC seeded intraluminally and normal human astrocytes seeded into the extra-capillary space (ECS). SIVmac251 was introduced intraluminally in the presence of polybrene, and infection and barrier formation were monitored. Infectious virus was assayed both by co-culture with indicator CEMx174 cells and by s-MAGI assay (Chackerian et al., 1995). In addition, samples for p27 ELISA were taken from both the lumen and the ECS. The results of these studies (Table 3) indicate that virus was consistently present thoughout the time course of infection for a total of 116 days.
In addition to virus production, lactate production by the BMEC as an indicator of cell metabolism was measured (Figure 5).
| Figure 5. Average rate of lactate production in SIV-infected vs. mock-infected DIV-BBB cartridges. |
The results show little or no difference in the rate of lactate produced between infected and mock-infected DIV-BBB cartridges. This observation is an important result demonstrating that infection of BMEC by SIV did not result in direct toxicity to the cells as measured by their metabolic rate.
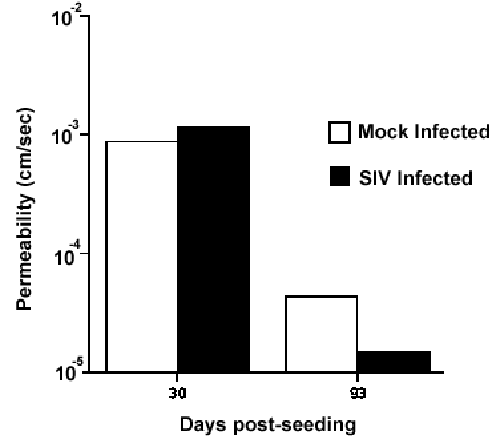
An additional important finding was that the cells of the DIV-BBB could be infected in the presence of a forming barrier (Figure 6).
| Figure 6. Permeability to sucrose in SIV-infected vs. mock-infected DIV-BBB cartridges. |
The establishment of the barrier, as measured by permeability to labeled sucrose, was not significantly decreased by viral infection.
Conclusions
Taken together, these preliminary results demonstrate that the DIV-BBB model described above demonstrates properties, which closely approximate the in vitro BBB. The mechanisms, which could be studied in this model, would also include mechanisms of viral persistence and activation, viral entry into the CNS through the BBB, and viral interference with either tight junction formation or with transporter function. Lastly, HIV infection of BMEC has been shown to induce adhesion molecules involved in the seeding of lymphomas (Moses et al., 1997). The DIV-BBB model will prove valuable in examining mechanisms involved in this process as well.
References
Bradbury, M. W. 1993. The Blood-Brain Barrier. Exper. Physiol 78:453-472. Medline
Chackerian, B., N. L. Haigwood, and J. Overbaugh. 1995. Characterization of a CD4-expressing macaque cell line that can detect virus after a single replication cycle and can be infected by diverse simian immunodeficiency virus isolates. Virology 213:386-394. Medline
Joo, F. 1985. The blood-brain barrier in vitro: Ten years of research on microvessels isolated from brain. Neurochem. Int. 7:1-5.
Joo, F. 1993. The blood-brain barrier in vitro: The second decade. Neurochem Intern. 23:499-521.
Mankowski, J. L., J. P. Spelman, H. G. Ressetar, J. D. Strandberg, J. Laterra, D. L. Carter, J. E. Clements, and M. C. Zink. 1994. Neurovirulent Simian Immunodeficiency Virus Replicates Productively in Endothelial Cells of the Central Nervous System In vivo and In vitro. J Virol 68:8202-8208. Medline
Moses, A. V., F E. Bloom, D. Pauza, and J. A. Nelson. 1993. Human Immunodeficiency Virus Infection of Human Brain Capillary Endothelial Cells Occurs via a CD4/Galactosylceramide-Independent Mechanism. Proc Natl Acad Sci 90:10474-10478. Medline
Moses, A. V, S. E. Williams, J. G. Strussenberg, M. L. Heneveld, R. A. Ruhl, A. C. Bakke, G. C. Bagby, and J. A. Nelson. 1997. HIV-1 induction of CD40 on endothelial cells promotes the outgrowth of AIDS-associated B-cell lymphomas. Nature Med 3:1242-1249. Medline
Ott, M. J., J. L. Olson, and B. J. Ballerman. 1995. Chronic in vitro flow promotes ultrastructural differentiation of endothelial cells. Endothelium 3:21-30.
Pardridge, W.M., D. Triguero, J. Yang and PA Cancilla. 1990. Comparison of in vitro and in vivo models of drug transcytosis through the blood-brain barrier. J. Pharmacol. Exp. Ther. 253:884-891. Medline
Petito, C. K., and K. S. Cash. 1992. Blood Brain Barrier Abnormalities in the Acquired Immunodeficiency Syndrome: Immunohistochemical Localization of Serum Proteins in Postmortem Brain. Ann Neurol 32:658-666. Medline
Rhodes, R. H. 1991. Evidence of Serum Protein Leakage Across the Blood Brain Barrier in the Acquired Immunodeficiency Syndrome. J Neuropathol Exp Neurol 50:171-183. Medline
Sasseville, V. G., W. Newman, S. J. Brodie, P. Hesterberg, D. Pauley, and D. J. Ringler. 1994. Monocyte Adhesion to Endothelium in Simian Immunodeficiency Virus-Induced AIDS Encephalitis is Mediated by Vascular Cell Adhesion Molecule-1/a4b1 Integrin Interactions. Am J Pathol 144:27-40. Medline
Stanness, K. A., E. Guatteo, and D. Janigro. 1996. A Dynamic Model of the Blood-Brain Barrier quot;In vitroquot; NeuroToxicology 17:481-496. Medline
Stanness, K. A., L. E. Westrum, E. Fornaciari, P. Mascagni, J. A. Nelson, S. G. Stenglein, T. Meyers, and D. Janigro. 1997. Morphologcal and Functional Characterization of an In vitro Blood-Brain Barrier Model. Brain Research, in press.
Wiley, C. A., R. D. Schrier, F. J. Denaro, J. A. Nelson, P. W. Lampert, and M. B. A.. Oldstone. 1986. Cellular localization of human immunodeficiency virus infection within the brains of acquired immune deficiency syndrome patients. Proc. Natl. Acad. Sci. USA. 83:7089-7093. Medline
| Copyright Information | Site map |