|
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Neutralizing antibodies (NAbs) were measured in an MT-2 cell-killing assay, using a stock of SHIV-89.6P expanded in human PBMCs as described (35). Detectable NAb titers were measured in 11 of 12 vaccinated monkeys by day 21 after challenge and in two of eight control monkeys (780 and KPE) by day 28 after challenge (Fig. 2B). All animals that developed detectable NAb responses had comparable peak titers.
Six of the eight control monkeys (all except 780 and KPE) demonstrated a rapid and profound depletion of their CD4+ T lymphocytes between days 7 and 21 after challenge, consistent with our previous experience with SHIV-89.6P infection (Fig. 3A) (33, 34). In contrast, all monkeys that received plasmid DNA vaccines plus IL-2/Ig protein or IL-2/Ig plasmid had complete preservation of their CD4+ T lymphocytes, with no evidence of declining CD4+ T cell counts by day 126 after challenge. At day 70 after challenge, a time by which setpoint of viral replication is reached in SHIV-89.6P-infected rhesus monkeys, a highly significant difference in peripheral blood CD4+ T cell counts was evident between the control monkeys and the monkeys that received the DNA vaccines plus IL-2/Ig protein or plasmid [P = 0.00006, by a two-sided Wilcoxon rank sum test (36)]. The monkeys that received the DNA vaccines alone also had detectable CD4+ T cell declines, but these were less dramatic than in the control monkeys (P = 0.2). In these animals, a marked but gradual CD4+ T cell decline was observed in monkey 811, and small CD4+ T cell declines were evident in monkeys 660 and 820.
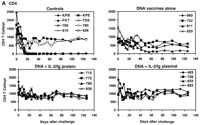
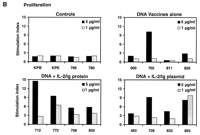
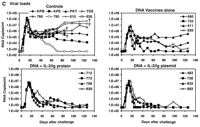
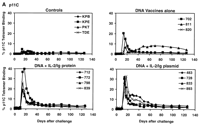
Fig. 3. Postchallenge CD4+ T lymphocyte counts, virus-specific lymphoproliferative responses, and viral loads. (A) CD4+ T lymphocyte counts in peripheral blood were determined by multiplying the total lymphocyte count by the percentage of CD3+CD4+ lymphocytes. (B) Lymphoproliferative responses to SIV Gag p28 protein at day 140 after challenge. Standard overnight thymidine incorporation assays were performed after incubation of PBMCs with antigen for 5 days in culture. Stimulation indices were calculated by: (counts per minute with antigen)/(counts per minute with media). (C) Plasma viral loads were determined at multiple time points after challenge by an ultrasensitive branched DNA amplification assay with a detection limit of 400 copies/ml (Bayer Diagnostics). Dagger (
To determine whether the challenged monkeys retained functional
virus-specific CD4+ T lymphocyte activity, we studied
their peripheral blood lymphoproliferative responses
after stimulation with purified SIV Gag p28 protein
(Intracel) and their CD4+ T cell IFN-![]() responses after stimulation with SIV Gag and HIV-1 Env
peptide pools (37).
At day 140 after challenge, all the surviving
control monkeys, including the monkeys with preserved
total CD4+ T lymphocyte counts (780 and KPE), had no
detectable lymphoproliferative responses to SIV Gag
(stimulation indices < 2) (Fig.
3B). In contrast, all the monkeys that received the
IL-2/Ig-augmented DNA vaccines had detectable
lymphoproliferative responses to SIV Gag, with
stimulation indices of 3.7 to 14.5. One of the
monkeys that received the DNA vaccines alone (monkey 702)
also had a detectable lymphoproliferative response. The
preservation of virus-specific CD4+ T
lymphocyte function in the vaccinated animals was confirmed
by intracellular IFN-
responses after stimulation with SIV Gag and HIV-1 Env
peptide pools (37).
At day 140 after challenge, all the surviving
control monkeys, including the monkeys with preserved
total CD4+ T lymphocyte counts (780 and KPE), had no
detectable lymphoproliferative responses to SIV Gag
(stimulation indices < 2) (Fig.
3B). In contrast, all the monkeys that received the
IL-2/Ig-augmented DNA vaccines had detectable
lymphoproliferative responses to SIV Gag, with
stimulation indices of 3.7 to 14.5. One of the
monkeys that received the DNA vaccines alone (monkey 702)
also had a detectable lymphoproliferative response. The
preservation of virus-specific CD4+ T
lymphocyte function in the vaccinated animals was confirmed
by intracellular IFN-![]() staining after stimulation with SIV Gag or HIV-1 Env
peptide pools (Table
2).
staining after stimulation with SIV Gag or HIV-1 Env
peptide pools (Table
2).
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Viremia and clinical disease progression. We next measured plasma viral loads in the monkeys by an ultrasensitive branched DNA amplification assay with a detection limit of 400 copies/ml (Bayer Diagnostics). The control monkeys had 1.0 × 107 to 1.8 × 108 copies/ml of virus at the time of peak viremia on day 14 after challenge (Fig. 3C). At day 70 after challenge, six of eight control monkeys had high setpoint viral loads of between 5.0 × 105 and 3.1 × 106 copies/ml. The other two control monkeys had lower setpoint viral loads: 4.6 × 104 copies/ml in monkey KPE and 4.4 × 102 copies/ml in monkey 780. In all the vaccinated monkeys, peak viremia was between 1.2 × 106 and 1.0 × 107 copies/ml, levels significantly lower than in the controls [P = 0.004 and P = 0.008 for monkeys receiving the DNA plus IL-2/Ig or the DNA vaccines alone, respectively, by two-sided Wilcoxon rank-sum tests (36)]. At day 70, setpoint viral loads in the monkeys receiving the IL-2/Ig-augmented DNA vaccines were also significantly lower than in the controls (P = 0.004), and a trend toward reduction in setpoint viral loads was evident in the animals that received the DNA vaccines alone (P = 0.1). At the majority of time points after setpoint, the monkeys that received the DNA vaccines plus IL-2/Ig plasmid had undetectable viremia (<400 copies/ml), although small and transient rises in viremia were seen in monkeys 728 and 893. In the monkeys that received IL-2/Ig protein, viremia was also generally controlled to under 103 copies/ml. The monkeys that received the DNA vaccines alone had a heterogeneous outcome, with high setpoint viremia in monkey 811, moderate setpoint viremia in monkeys 660 and 820, and controlled viremia in monkey 702.
Compared with the control monkeys, the animals that received the
DNA vaccines alone had a 1.9 log reduction in geometric
mean viral load after setpoint. The magnitude of this
reduction is similar to that seen in previous studies
from our laboratory assessing the efficacy of an SIV Gag
DNA vaccine (12)
or a recombinant MVA-Gag/Pol vaccine (38)
in conjunction with SIVsm E660 challenges. The monkeys
that received the DNA vaccines plus IL-2/Ig protein or
plasmid in the present study had >2.7 and >3.0 log
reductions, respectively, in geometric mean viral load after
setpoint compared with the control monkeys. In fact, the
control of setpoint viremia observed in the monkeys that
received the DNA vaccines plus IL-2/Ig plasmid was
similar in magnitude to the control of SHIV-89.6P
setpoint viremia achieved by immunization with a live,
attenuated SIVmac239![]() 3
vaccine (39),
although the heterologous challenge in that study limits
the comparability of results between these reports.
3
vaccine (39),
although the heterologous challenge in that study limits
the comparability of results between these reports.
The pathogenicity of the SHIV-89.6P challenge virus was confirmed by the rapid clinical disease progression in the control monkeys that developed low CD4+ T lymphocyte counts and high viremia. Seven of the eight control monkeys (all except monkey 780) developed significant clinical disease, and four of these eight monkeys died by day 140 after challenge (Fig. 4 and Table 3). In contrast, all monkeys that received the cytokine-augmented DNA vaccines remained healthy with no documented clinical events or mortality. The prevention of clinical disease in the vaccinated monkeys as compared with the controls was highly significant (P = 0.001, by a two-sided Fisher exact test), and a trend was observed for preventing mortality (P = 0.08). The monkeys that received the DNA vaccines alone had an intermediate clinical outcome, with monkeys 811 and 820 showing signs of clinical disease (P = 0.24 compared with controls).
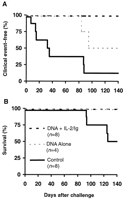 Fig. 4. Postchallenge clinical events
and mortality in the monkeys that received the cytokine-augmented
DNA vaccines, the DNA vaccines alone, or the sham control vaccine.
(A) Percentage of monkeys free from clinical events directly
attributable to the SHIV-89.6P infection or the subsequent
immunodeficiency. (B) Survival curve.
Fig. 4. Postchallenge clinical events
and mortality in the monkeys that received the cytokine-augmented
DNA vaccines, the DNA vaccines alone, or the sham control vaccine.
(A) Percentage of monkeys free from clinical events directly
attributable to the SHIV-89.6P infection or the subsequent
immunodeficiency. (B) Survival curve.
| ||||||||||||||||||||||||||||||||||||||||
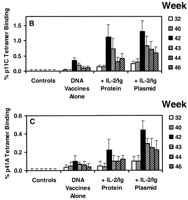
Immune correlates of protection. Scatter plots of data from the Mamu-A*01-positive animals in this study (Fig. 5, A and B) demonstrate significant correlations of prechallenge vaccine-elicited plateau-phase p11C-specific CTL responses determined by tetramer staining, with postchallenge peak p11C-specific CTL responses (P = 0.007, by a two-sided Spearman rank correlation test) and setpoint viral loads (P = 0.04). Levels of peak vaccine-elicited CTLs were less predictive of outcome than those of plateau-phase vaccine-elicited CTLs, the latter presumably reflecting the memory cell population. The asymptotic appearances of the plotted data suggest that a level of vaccine-elicited plateau-phase p11C-specific CTLs may exist (approximately 0.3 to 0.5% of CD3+CD8+ T cells) above which little additional benefit is discernible after challenge. Similar significant correlations were observed between plateau-phase p41A-specific CTL responses before challenge and peak p41A-specific CTL responses and setpoint viral loads after challenge (31). These results strongly suggest that the improved outcome in animals receiving the cytokine-augmented DNA vaccines resulted from the augmented vaccine-elicited CTL responses in these animals.
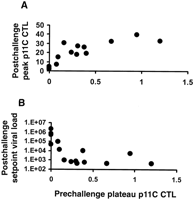 Fig. 5. Correlation of prechallenge
vaccine-elicited plateau-phase p11C-specific CTL responses as
determined by tetramer staining with (A) postchallenge peak
p11C-specific CTL responses and (B) setpoint viral loads in
the Mamu-A*01-positive rhesus monkeys.
Fig. 5. Correlation of prechallenge
vaccine-elicited plateau-phase p11C-specific CTL responses as
determined by tetramer staining with (A) postchallenge peak
p11C-specific CTL responses and (B) setpoint viral loads in
the Mamu-A*01-positive rhesus monkeys. No neutralizing antibody (NAb) responses were detected in the vaccinated monkeys prior to challenge, and comparable peak NAb titers developed in the vaccinated monkeys and the two control monkeys with preserved CD4+ T lymphocyte counts. The other six control monkeys that rapidly lost their CD4+ T lymphocytes would not be expected to generate significant NAb responses (34). Whereas peak CTL responses occurred on day 14 after challenge, coincident with the peak of primary viremia, NAb responses were generally first detectable on day 21 when viremia had already been partially controlled. The initial control of primary viremia in these monkeys therefore appears to be primarily due to CTL activity, as has previously been observed in HIV-infected humans and SIV-infected monkeys (40-42). However, NAbs may also have played a significant role in the control of acute or chronic viremia. In addition, it is possible that IL-2/Ig administration may have modulated nonspecific innate immune functions in these animals and contributed to the containment of virus replication.
Conclusions. In summary, after the SHIV-89.6P challenge, 75% of control animals developed weak immune responses, rapid and profound loss of CD4+ T cells, high viral loads, and rapid disease progression. The Mamu-A*01-positive and the Mamu-A*01-negative control monkeys had similar outcomes, and thus this particular MHC class I allele conferred no particular protective or detrimental effect. It is unclear why monkeys 780 and KPE had more favorable outcomes than the other control animals. This may reflect higher virus-specific immune responses or other protective host factors in these animals. The monkeys that received the cytokine-augmented DNA vaccines, in contrast, had uniformly good outcomes with potent immune responses, preserved CD4+ T cell counts, low to undetectable viral loads, and no evidence of clinical disease. Moreover, these animals also demonstrated preserved virus-specific CD4+ T cell responses, which may confer long-term clinical benefits (43). The monkeys that received the DNA vaccines alone had intermediate and heterogeneous outcomes. Their postchallenge CTL levels reflected both the levels of prechallenge CTLs as well as the levels of viremia driving these responses.
The administration of IL-2/Ig protein or IL-2/Ig plasmid during initial DNA vaccine priming led to augmented immune responses that were capable of controlling viremia and preventing immunodeficiency, clinical disease, and death following a homologous pathogenic SHIV-89.6P challenge in rhesus monkeys. These results raise the possibility that viral replication may also be reduced in humans who have been similarly vaccinated and subsequently infected with HIV-1. As a consequence, such individuals might manifest decreased disease burden and HIV-1 transmission rates (44, 45). Moreover, this strategy of augmenting vaccine-elicited immune responses by cytokine administration should be readily applicable to other vaccine modalities and for other immunotherapeutic purposes.
REFERENCES AND NOTES
- A. McMichael, Cell 93, 673 (1998) [ISI] [Medline].
- J. E. Schmitz, et al., Science 283, 857 (1999) [ISI] [Abstract/Full Text].
- X. Jin, et al., J. Exp. Med. 189, 991 (1999) [ISI] [Abstract/Full Text].
- L. Musey, et al., N. Engl. J. Med. 337, 1267 (1997) [ISI] [Medline].
- G. S. Ogg, et al., Science 279, 2103 (1998) [ISI] [Abstract/Full Text].
- J. A. Wolff, et al., Science 247, 1465 (1990) [ISI] [Medline].
- D. C. Tang, M. Devit, S. A. Johnson, Nature 356, 152 (1992) [ISI] [Medline].
- J. B. Ulmer, et al., Science 259, 1745 (1993) [ISI] [Medline].
- N. L. Letvin, et al., Proc. Natl. Acad. Sci. U.S.A. 94, 9378 (1997) [ISI] [Abstract/Full Text].
- J. D. Boyer, et al., Nature Med. 3, 526 (1997) [ISI] [Medline].
- S. Lu, et al., J. Virol. 70, 3978 (1996) [ISI] [Abstract].
- M. A. Egan, et al., J. Virol. 74, 7485 (2000) [ISI] [Abstract/Full Text].
- H. L. Robinson, et al., Nature Med. 5, 526 (1999) [ISI] [Medline].
- S. J. Kent, et al., J. Virol. 72, 10180 (1998) [ISI] [Abstract/Full Text].
- T. Hanke, et al., J. Virol. 73, 7524 (1999) [ISI] [Abstract/Full Text].
- T. M. Allen, et al., J. Immunol. 164, 4968 (2000) [ISI] [Abstract/Full Text].
- M. A. Geissler, A. Gesien, K. Tokushige, J. R. Wands, J. Immunol. 158, 1231 (1997) [ISI] [Abstract].
- Y.-H. Chow, W.-L. Huang, W.-K. Chi, Y.-D. Chu, M.-H. Tao, J. Virol. 71, 169 (1997) [ISI] [Abstract].
- K.-Q. Xin, et al., Immunology 94, 438 (1998) [ISI] [Medline].
- N. F. Landolfi, J. Immunol. 146, 915 (1991) [ISI] [Abstract].
- P. Nickerson,
et al., Transplant. Immunol. 4, 81 (1996)
[Medline]. - D. H. Barouch, et al., J. Immunol. 161, 1875 (1998) [ISI] [Abstract/Full Text].
- D. H. Barouch, et al., Proc. Natl. Acad. Sci. U.S.A. 97, 4192 (2000) [ISI] [Abstract/Full Text].
- Large-scale preparations of plasmids were carried out by alkaline lysis and CsCl gradient banding. Rhesus monkeys were vaccinated by intramuscular (i.m.) injections of either 5 mg of HIV-1 89.6P env DNA plus 5 mg of SIVmac239 gag DNA or 10 mg of sham DNA in sterile saline without adjuvant. Half the dose was delivered to each quadriceps muscle in a 0.5-ml volume using a needle-free Biojector apparatus (Bioject). Monkeys were vaccinated at weeks 0, 4, 8, and 40. At weeks 0 and 4, monkeys that received the DNA vaccines alone also received 5 mg of sham DNA 2 days following vaccination (660, 702, 811, 820); monkeys that received human IL-2/Ig protein received 0.5 mg/day affinity-purified protein in two divided doses i.m. on days 1 through 14 following vaccination (712, 772, 798, 839); and monkeys that received human IL-2/Ig plasmid received 5 mg of IL-2/Ig DNA 2 days following vaccination (483, 728, 833, 893). All the vaccinated monkeys except 660 expressed the Mamu-A*01 class I allele. Of the control monkeys, four were Mamu-A*01-negative (766, 780, 810, 826). The four control monkeys that were Mamu-A*01-positive (KPB, KPE, PKT, TDE) were added to the study prior to the week 40 boost immunization. All the animals used in this study were maintained in accordance with the guidelines of the Committee on Animals for Harvard Medical School, the animal care protocols of Southern Research Institute, and the Guide for the Care and Use of Laboratory Animals (National Research Council, National Academy Press, Washington, DC, 1996). After challenge, the immunologic assays, viral load measurements, and clinical care of the animals were performed blinded to the immunizations each monkey received.
- Single-letter abbreviations for the amino acid residues are as follows: A, Ala; C, Cys; D, Asp; G, Gly; I, Ile; L, Leu; M, Met; N, Asn; P, Pro; Q, Gln; R, Arg; S, Ser; T, Thr; V, Val; and Y, Tyr.
- M. D. Miller, H. Yamamoto, A. H. Hughes, D. I. Watkins, N. L. Letvin, J. Immunol. 147, 320 (1991) [ISI] [Abstract].
- T. M. Allen, et al., J. Immunol. 160, 6062 (1998) [ISI] [Medline]. We have simplified previous terminology for the Mamu-A*01-restricted immunodominant SIV Gag epitope from p11C,C-M to p11C for convenience.
- M. A. Egan, et al., J. Virol. 73, 5466 (1999) [ISI] [Abstract/Full Text].
- J. D. Altman, et al., Science 274, 94 (1996) [ISI] [Abstract/Full Text].
- M. J. Kuroda, et al., J. Exp. Med. 187, 1373 (1998) [ISI] [Abstract/Full Text]. Briefly, 1 µg of phycoerythrin-labeled tetrameric Mamu-A*01/peptide complexes was used in conjunction with fluorescein isothiocyanate (FITC)-labeled anti-human CD8
 (Leu2a; Becton-Dickinson), ECD-labeled anti-human CD8
(Leu2a; Becton-Dickinson), ECD-labeled anti-human CD8 (2ST8-5H7; Beckman Coulter), and APC-labeled anti-rhesus monkey
CD3 (FN18; Biosource) monoclonal antibodies to stain
peptide-specific CD8+ T cells. Whole blood
(100 µl) from the vaccinated monkeys was directly stained
with these reagents, then lysed, washed, and fixed. Samples were
analyzed by four-color flow cytometry on a Coulter EPICS Elite ESP
system, and gated CD3+CD8+ T cells were
examined for staining with tetrameric Mamu-A*01/p11C,
Mamu-A*01/p41A, or Mamu-A*01/p68A complexes.
(2ST8-5H7; Beckman Coulter), and APC-labeled anti-rhesus monkey
CD3 (FN18; Biosource) monoclonal antibodies to stain
peptide-specific CD8+ T cells. Whole blood
(100 µl) from the vaccinated monkeys was directly stained
with these reagents, then lysed, washed, and fixed. Samples were
analyzed by four-color flow cytometry on a Coulter EPICS Elite ESP
system, and gated CD3+CD8+ T cells were
examined for staining with tetrameric Mamu-A*01/p11C,
Mamu-A*01/p41A, or Mamu-A*01/p68A complexes.
- D. H. Barouch et al., data not shown.
- K. A. Reimann, et al., J. Virol. 70, 3198 (1996) [ISI] [Abstract].
- K. A. Reimann, et al., J. Virol. 70, 6922 (1996) [ISI] [Abstract].
- K. A. Reimann, et al., Virology 256, 15 (1999) [ISI] [Medline].
- J. M. Crawford, et al., J. Virol. 73, 10199 (1999) [ISI] [Abstract/Full Text].
- Statistical analysis was performed with GraphPad Prism, version 2.01 (GraphPad Software, Inc., 1996). CD4+ T lymphocyte counts and viral loads were compared between groups by two-sided Wilcoxon rank-sum tests with Bonferroni adjustments of P values to account for the two major comparisons of each endpoint. Day 70 setpoint values were chosen in order to analyze a complete data set prior to death of any animals. Differences in clinical events and mortality were analyzed by two-sided Fisher exact tests. Correlations of prechallenge vaccine-elicited CTLs and postchallenge peak CTLs or setpoint viral loads were assessed in the vaccinated monkeys by two-sided Spearman rank correlation tests. In all cases, P < 0.05 was considered significant.
- Intracellular cytokine staining assays were performed as follows. We placed 2 × 106 PBMCs in 17 mm by 100 mm polypropylene tubes containing 1 ml of supplemented RPMI medium, to which 1 µg of each costimulatory antibody (anti-CD28 and anti-CD49d, Becton-Dickinson) was added. Peptide pools were added at a final concentration of 2 µg/ml. Culture tubes were incubated at a 5° slant at 37°C in a humidified 5% CO2 incubator for 16 hours. Brefeldin A (Sigma) was added for the last 15 hours at a final concentration of 10 µg/ml. Cells were stained with anti-CD3-APC (FN18, Biosource), anti-CD4-PE (OKT4, Ortho Diagnostics), and anti-CD8-PerCP (SK1, Becton-Dickinson) for 30 min and washed with phosphate-buffered saline containing 1% fetal bovine serum. Cells were permeabilized with FACS Permeabilization Buffer (Becton-Dickinson), washed, and stained with anti-IFN-
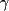 -FITC
(MD1, Biosource) for 30 min. Samples were fixed in 1%
formaldehyde and analyzed on a FACSCalibur flow cytometer
(Becton-Dickinson). In order to measure responses of
CD4+ T lymphocytes, 30,000 gated
CD3+CD4+ lymphocytes were analyzed for
intracellular IFN-
staining events using CellQuest software (Becton-Dickinson).
-FITC
(MD1, Biosource) for 30 min. Samples were fixed in 1%
formaldehyde and analyzed on a FACSCalibur flow cytometer
(Becton-Dickinson). In order to measure responses of
CD4+ T lymphocytes, 30,000 gated
CD3+CD4+ lymphocytes were analyzed for
intracellular IFN-
staining events using CellQuest software (Becton-Dickinson).
- A. Seth, et al., J. Virol. 74, 2502 (2000) [ISI] [Abstract/Full Text].
- M. S. Wyand, et al., J. Virol. 73, 8356 (1999) [ISI] [Abstract/Full Text].
- R. A. Koup, et al., J. Virol. 68, 4650 (1994) [ISI] [Abstract].
- G. Pantaleo, et al., Nature 370, 463 (1994) [ISI] [Medline].
- Z. W. Chen, et al., J. Exp. Med. 182, 21 (1995) [ISI] [Abstract].
- E. S. Rosenberg, et al., Science 278, 1447 (1997) [ISI] [Abstract/Full Text].
- P. M. Garcia, et al., N. Engl. J. Med. 341, 394 (1999) [ISI] [Medline].
- T. C. Quinn, et al., N. Engl. J. Med. 342, 921 (2000) [ISI] [Medline].
- We acknowledge support from NIH grants CA-50139 (N.L.L), AI-85343 (N.L.L. and D.C.M.), AI-65301 (M.G.L.), AI/GF-41521 (T.B.S.), and AI-42298 (T.B.S.). We are grateful to N. Miller, F. Vogel, M. Forman, K. Reimann, W. Lin, A. Miura, R. Kuhnkuhn, C. Lord, J. Frost, T. Steenbeke, C. Crabbs, J. Yalley-Ogunro, N. Persaud, L. Zhu, and J. Joyce for generous advice, assistance, and reagents.
Related articles in Science:
- Helminthic Infection and HIV Vaccine
Trials.
- Miles B. Markus, John E. Fincham;, Dan H. Barouch, and Norman
L. Letvin
Science 2001 291: 46-47. (in Letters)[Full Text] - AIDS:
Enhanced: Preventing AIDS But Not HIV-1 Infection with a DNA Vaccine.
- Xuefei Shen and Robert F. Siliciano
Science 2000 290: 463-465. (in Perspectives)[Summary] [Full Text]
Volume 290, Number 5491, Issue of 20 Oct 2000, pp. 486-492.
Copyright © 2000 by The American Association for the Advancement of Science.